Morfologia externa das plantas angiospermas
O corpo da maioria das plantas angiospermas é dividido em duas partes principais, uma localizada sob o solo, constituída pelas raízes, e outra área constituída pelo caule, folhas, flores e frutos. As células das raízes, assim com as células de muito caules, não fazem fotossíntese e por isso dependem do alimento produzido nas células das folhas. O caule, folhas, flores e frutos, por sua vez, dependem da água e dos sais minerais absorvidos pelas raízes.
A raiz
Quase sempre a raiz é originada a partir da radícula do embrião, localizado na semente.
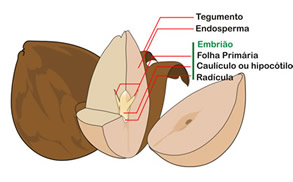
Partes das semente
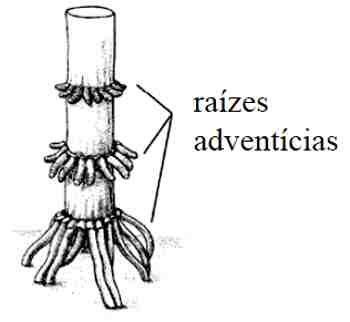
A partir dela surgem ramos secundários. No entanto, é freqüente surgirem raízes a partir de caules e mesmo de folhas. Essas raízes conhecidas como adventícias (do latim advena = que vem de fora, que nasce fora do lugar habitual), são comuns, por exemplo, na base de um pé de milho.
As raízes distribuem-se amplamente pelo solo, mas há algumas plantas que possuem raízes aéreas, comuns nas trepadeiras, bromélias, orquídeas, enquanto outras possuem raízes submersas, como os aguapés, comuns em represas.
 | |  |
Raiz aérea
| |
Raiz aquática
|
Temos dois tipos básicos de sistema radicular: o pivotante, em que há uma raiz principal, e ofasciculado, em que os ramos radiculares são equivalentes em tamanho e aparência, não apresentando uma raiz principal.
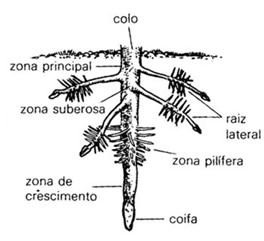
Partes da raiz
A extremidade de uma raiz é envolta por um capuz de células denominado coifa, cuja função é proteger omeristema radicular, um tecido em que as células estão se multiplicando ativamente por mitose. É no meristema que são produzidos as novas células da raiz, o que possibilita o seu crescimento.
Logo após a extremidade, localiza-se a região onde as células surgidas por mitose crescem. Nessa região denominada zona de distensão ou de alongamento celular, a raiz apresenta a maior taxa de crescimento.
Após a zona de distensão situa-se a zona pilífera da raiz, que se caracteriza por apresentar células epidérmicas dotadas de projeções citoplasmáticas finas e alongadas, os pêlos absorventes. É através desses pelos que a raiz absorve a maior parte da água e dos sais minerais de que precisa.
Já a região de ramos secundários é aquela que se nota o brotamento de novas raízes que surgem de regiões internas da raiz principal.
| |  |
Tipos de Raizes
A principal função da raiz é a absorção dos nutrientes minerais, sendo que, no solo, também é responsável pela fixação do vegetal ao substrato. Alguns tipos de raízes, no entanto, também desempenham outras funções:
Raizes tuberosas, como as da mandioca, da batata-doce e do nabo armazenam reservas alimentares, principalmente na forma de grãos de amido, utilizadas durante a floração e a produção de frutos pela planta. Os agricultores colhem essas raízes antes da planta tenha chance de consumir as reservas armazenadas, utilizando-as na alimentação humana e de animais.
Raízes respiratórias ou pneumatóforos são adaptadas a realização de trocas gasosas com o ambiente. Esse tipo de raiz é encontrado em plantas como a Avicena tomentosa, que vive no solo encharcado e pobre em gás oxigênio nos manguezais. As raízes principais dessa planta crescem rente à superfície do solo e, de espaço em espaço, apresentam pneumatóforos, que crescem para cima, perpendicularmente ao solo. Durante a maré vazante os pneumatóforos ficam expostos e pode realizar trocas de gases com o ar.
Raízes-suportes, também chamadas raízes-escoras, aumentam a base de fixação da planta ao solo. Algumas espécies de árvores possuem raízes tubulares, em forma de pranchas verticais, que aumentam a estabilidade da planta e fornecem maior superfície para respiração do sistema radicular.
Raízes aéreas são características de plantas epífetas, isto é, que vivem sobre outras plantas sem parasitá-las. Essas raízes podem atingir vários metros de comprimento antes de alcançar o solo, constituindo os cipós.
Raízes sugadoras são adaptadas à extração de alimentos de plantas hospedeiras, sendo características de plantas parasitas, como o cipó-chumbo e a erva-de-passarinho. As raízes sugadoras possuem um órgão de fixação, chamado apreensório, do qual partem finas projeções denominadas haustórios. Oshaustórios penetram na planta hospedeira até atingir os vasos condutores de seiva, de onde extraem água e nutrientes de que a planta parasita necessita para sobreviver.
No caso de a planta ser hemiparasita, a exemplo da erva-de-passarinho (é clorofilada, e portanto autótrofa), somente a seiva bruta (água e minerais), que transita pelos vasos lenhosos do xilema, é retirada da planta hospedeira.
Caule
As funções do caule
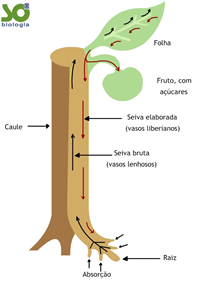 | |
O caule realiza a integração de raízes e folhas, tanto do ponto de vista estrutural como funcional. Em outras palavras, além de constituir a estrutura física onde se inserem raízes e folhas, o caule desempenha as funções de condução de água e sais minerais das raízes para as folhas, e de condução de matéria orgânica das folhas para as raízes.
Caules jovens têm células clorofiladas e são revestidos por uma epiderme uniestratificada, isto é, formada por uma única camada (estrato) de células. Plantas que apresentam pequeno crescimento em espessura, como as gramíneas, por exemplo, também apresentam caules revestidos pela epiderme e esta pode ainda apresentar sobre si, externamente, uma cutícula protetora.
Já em plantas que crescem muito em espessura, transformando-se em arbustos ou árvores, a epiderme é substituída por um revestimento complexo, formado por vários tecidos. O tecido mais externo é formado por células mortas, que conferem o aspecto áspero e opaco aos troncos das árvores. Esse revestimento multitecidual, denominado periderme, acompanha o crescimento em espessura dos troncos.
|
Os caules são, em geral, estruturas aéreas, que crescem verticalmente em relação ao solo. Existem, no entanto, caules que crescem horizontalmente, muitas vezes, subterraneamente.
Caules subterrâneos podem ser distinguidos de raízes porque apresentam gemas ou botões vegetativos, a partir dos quais podem se desenvolver ramos e folhas.
Gemas
As gemas caulinares são formadas por grupos de células meristemáticas, capazes de se multiplicar ativamente por mitose. Um conjunto de células meristemáticas forma um meristema, motivo pela qual as gemas caulinares também são chamadas meristemas caulinares.
No ápice do caule (e de cada ramo) existe sempre uma gema (ou meristema) apical, que permite o crescimento em extensão graças à multiplicação das células meristemáticas. À medida que o caule cresce diferenciam-se lateralmente, regiões onde surgem folhas e gemas axilares (ou laterais). As regiões onde se inserem as folhas e as gemas são denominadas nós e os espaços entre os nós são chamados entrenós.
As gemas axilares são meristemas localizados no caule, junto ao ângulo formado entre a folha e o ramo, que os botânicos denominaram “axila” foliar. As gemas axilares permanecem inativas durante certo período, denominado dormência após o qual podem entrar em atividade, originando ramos laterais.
Tipos de caules
Troncos são caules robustos, desenvolvidos na parte inferior e ramificados no ápice. São encontrados na maioria das árvores e arbustos do grupo das dicotiledôneas.
Estipes são caules geralmente não ramificados, que apresentam em seu ápice um tufo de folhas. São típicos das palmeiras.
Colmos são caules não-ramificados que se distinguem dos estipes por apresentarem, em toda a sua extensão, divisão nítida em gomos. Os gomos dos colmos podem ser ocos como no bambu, ou cheios como no milho ou na cana-de-açúcar.
Caules trepadores estão presentes em plantas trepadeiras e crescem enrolados sobre diversos tipos de suporte. Esse tipo de caule representa uma adaptação à obtenção de locais mais iluminados, em que há mais luz para a fotossíntese.
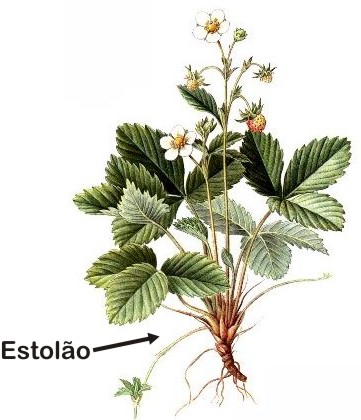
Estolão ou estolho é um tipo de caule que cresce paralelamente ao chão, produzindo gemas de espaço em espaço. Essas gema podem formar raízes e folhas e originar novas plantas.
Rizomas são caules subterrâneos que acumulam substâncias nutritivas. Em alguns rizomas ocorre acúmulo de material nutritivo em certas regiões, formando tubérculos. Rizomas podem ser distinguidos de raízes pelo fato de apresentarem gemas laterais. O gengibre, usado como tempero na cozinha oriental, é um caule tipo rizoma.
Na bananeira, o caule é um rizoma e a parte aérea é constituída exclusivamente por folhas. Um única vez na vida de uma bananeira um ramo caulinar cresce para fora do solo, dentro do conjunto de folhas, e forma em seu ápice uma inflorescência que se transforma em um cacho com várias pencas de bananas.
A batata-inglesa possui um caule subterrâneo que forma tubérculos, as batatas, um dos alimentos mais consumidos no mundo.

Bulbos são estruturas complexas formadas pelo caule e por folhas modificadas. Os bulbos costumam ser classificados em três tipos: tunicado, escamoso e cheio.
O exemplo clássico de bulbo tunicado é a cebola, cuja porção central, chamada prato, é pouco desenvolvida. Da parte superior do prato partem folhas modificadas, muito ricas em substâncias nutritivas: são os catafilos, que formam a cabeça da cebola. Da porção inferior do prato partem as raízes.
O bulbo escamoso difere do tunicado pelo fato dos catafilos se disporem como escamas parcialmente sobrepostas. Esse tipo de bulbo é encontrado no lírio.
No caso do bulbo cheio, as escamas são menos numerosas e revestem o bulbo como se fosse uma casca. Bulbos cheios estão presentes na palma.

Cladódios são caules modificados, adaptados à realização de fotossíntese. As plantas que os possuem perderam as folhas no curso da evolução, geralmente como adaptação a regiões de clima seco. A ausência de folhas permite à planta economizar parte da água que será perdida por evaporação.
Gavinhas são ramos modificados que servem para a fixação de plantas trepadeiras. Ao encontrar um substrato adequado as gavinhas crescem enrolando-se sobre ele.
Espinhos são ramos curtos, resistentes e com ponta afiada, cuja função é proteger a planta, afastando dela animais que poderiam danificá-la.Os espinhos tanto podem surgir por modificações de folhas, como nas cactáceas, como se originar do caule. Nesse caso forma-se nas axilas das folas, a partir de uma gema axilar, como ocorre nos limoeiros e laranjeiras.
Nas roseiras não há espinhos verdadeiros e sim acúleos, estruturas afiadas originadas da epiderme, o que explica serem facilmente destacáveis da planta, ao contrário dos espinhos.
A folha: local da fotossíntese
De formato extremamente variável, uma folha completa é formada por um “cabinho”, o pecíolo, e uma superfície achatada dotada de duas faces, o limbo percorrido pelas nervuras. A principal função da folha é servir como local em que é realizada a fotossíntese. Em algumas plantas, existem folhas modificadas e que exercem funções especializadas, como as folhas aprisionadoras de insetos das plantas insetívoras, e os espinhos dos cactos.
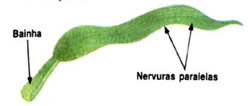
Uma folha é sempre originada a partir de um gema lateral do caule. Existem dois tipos básicos de folhas quanto ao tipo de nervura que apresentam: as paralelinérveas, típicas das monocotiledôneas, e asreticulinérveas, comuns em eudicotiledôneas.
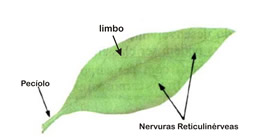
Eudicotiledôneas são uma das duas principais classes de angiospermas; inicialmente contidas dentro do grupo das dicotiledôneas, que foi desmembrado por não ser monofilético. O prefixo eu significaverdadeiro, portanto este termo designaria as plantas que realmente apresentam dois cotilédones. Esse grupo difere-se do antigo dicotiledônea por apresentar somente plantas que apresentem grão de pólen triaperturado, característica derivada de um ancestral comum, que torna o grupo monofilético
|
Algumas estruturas foliares especiais
Em algumas plantas, principalmente monocotiledôneas, não há um tecido propriamente dito, mas um estrutura conhecida pelo nome de bainha, que serve de elemento de ligação da folha à planta. É o caso, por exemplo, da folha de milho. Já em eudicotiledôneas, próximas aos pecíolos existem estruturas de formatos diversos – podem ser pontiagudas, laminares ou com a forma de espinhos – conhecidas porestípulas.
O formato e a cor das folhas são muito variáveis e algumas delas chamam a atenção por sua estrutura peculiar. É o caso por exemplo, das folhas modificadas presentes em plantas carnívoras, cuja adaptação auxilia na captura de insetos. Também é especialmente interessante a coloração de certasbrácteas, pequenas folhas modificadas na base das flores, apresentam: de tão coloridas, elas atuam como importante elemento para atração dos insetos.
Dionaea Muscipula- Planta carnívora
Plantas descíduas e abscisão foliar
 | |
Em muitas espécies de angiospermas, principalmente nas adaptadas a regiões temperadas, as folhas caem no outono e renascem na primavera. Plantas que perdem as folhas em determinada estação do ano são chamadas decíduas oucaducifólias. Plantas que não perdem as folhas são chamadas de perenes.
A queda das folhas no outono é interpretada como umaadaptação ao frio intenso e à neve. Em vez de ter as folhas lesadas pelo frio do inverno, a planta as derruba “deliberadamente” no outono, em um processo por ela controlado.
|
A queda das folhas ocorre por meio de um processo chamado abscisão foliar. Inicialmente forma-se um tecido cicatricial na região do pecíolo que une a folha ao caule, o tecido de abscisão, que interrompe gradativamente a passagem de água e nutrientes minerais do caule para a folha. A planta, assim, perde as folhas com o mínimo de prejuízo e reduz a atividade metabólica durante todo o inverno. Na primavera, surgem novos primórdios foliares junto às gemas dormentes, que logo se desenvolvem em folhas.
Classificação das folhas
As folhas podem ser classificadas de diversas maneiras: de acordo com a sua disposição no caule, a forma do limbo, a forma da borda etc.
Filotaxia
Filotaxia é o modo como as folhas estão arranjadas no caule. Existem três tipos básicos de filotaxia:oposta, verticilada e alternada.
A filotaxia é oposta quando existem duas folhas por nó, inseridas em regiões opostas. Quando três ou mais folhas inserem-se no mesmo nó, a filotaxia é chamada verticilada. Quando as folhas se inserem em regiões ligeiramente deslocadas entre si, em nós sucessivos, descrevendo uma hélice, a filotaxia é chamada alternada.
Folha oposta Folha verticilada
Tipos de limbo
O limbo pode ser simples (não-dividido) ou composto, dividido em dois, três ou mais folíolos. Caso os folíolos de um limbo composto partam todos de um mesmo ponto do pecíolo, dispondo-se como os dedos de uma mão, a folha é chamada de palmada.
Quando os folíolos de dispõem ao longo do pecíolo, a folha é chamada de penada. As folhas penadas podem terminar em um único folíolo, sendo chamadas imparipenadas, ou em dois folíolos, sendo chamadas paripenadas.
A forma e o tipo de borda do limbo são outras características utilizadas na classificação de folhas.
Flor
A flor é o órgão reprodutivo das plantas angiospermas. Flores que apresentam órgãos reprodutores de ambos os sexos, masculino e feminino, são chamadas de hermafroditas (ou monóica). Já as flores que apresentam órgãos reprodutores de apenas um dos sexos (masculino ou feminino) são chamadas dedióica.
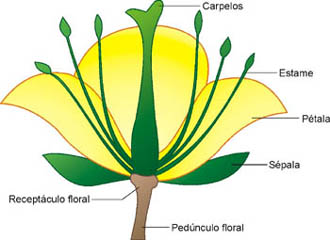
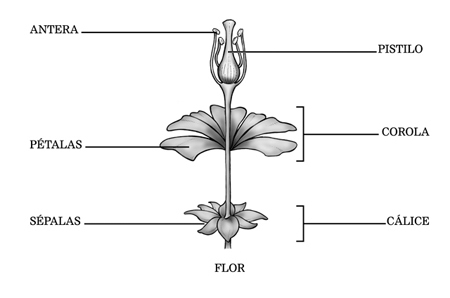
Uma flor hermafrodita é geralmente constituída por quatro conjuntos de folhas modificadas, os verticilos florais. Os verticilos se inserem em um ramos especializado, denominado receptáculo floral. Os quatro verticilos florais são o cálice, constituído pelas sépalas, a corola, constituída pelas pétalas, o androceu, constituído pelos estames, e o gineceu, constituído pelos carpelos.
Flores completas e incompletas
Uma flor que apresenta os quatro verticilos florais, ou seja, cálice, corola, androceu egineceu, é uma flor completa. Quando falta um ou mias desses componentes a flor é chamadaincompleta.
Cálice, corola e perianto
As sépalas são geralmente verdes e lembram folhas. São as partes mais externas da flor e a sua função é cobrir e proteger o botão floral antes dele se abrir. O conjunto de sépalas forma o cálice floral.
Pétalas são estruturas geralmente coloridas e delicadas e se localizam internamente às sépalas. O conjunto de pétalas forma a corola.
O conjunto formado pelos dois verticilos florais mais externos, o cálice e a corola, é denominado perianto(do grego Peri, em torno, e anthos, flor).
Estames
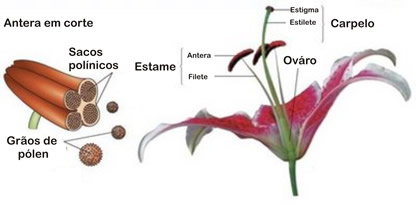
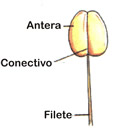
Estames são folhas modificadas, onde se formam os gametas masculinos da flor. O conjunto de estames forma o androceu (do grego andros, homem, masculino). Um estame geralmente apresenta uma parte alongada, o filete, e uma parte terminal dilatada, a antera.
O interior da antera é geralmente dividido em quatro cavidades, dentro das quais se formam os grãos de pólen. No interior de cada grão de pólen forma-se dois gametas masculinos, denominados núcleos espermáticos. Quando a flor está madura, as anteras se abrem e libertam os grãos de pólen.
| |  |
Carpelos
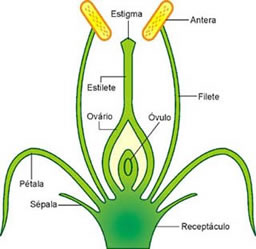
Carpelos são folhas modificadas, em que se formam os gametas feminios da flor. Um ou mais carpelos formam uma estrutura em forma de vaso, o pistilo. Este apresenta uma região basal dilatada, o ovário,do qual parte um tubo, o estilete, que termina em uma região dilatada, o estigma. O conjunto de pistilos de uma flor constitui o gineceu (do grego gyncos, mulher, feminino).
O pistilo pode ser constituído por um, dois ou mais carpelos, dependendo do tipo de flor. Em geral, o número de câmaras internas que o ovário apresenta corresponde ao número de carpelos que se fundiram para formá-lo. No interior do ovário formam-se um ou mais óvulos.
Os óvulos vegetais são estruturas complexas, constituídas por muitas células. Nisso os óvulos vegetais diferem dos óvulos animais, que são estruturas unicelulares.
No interior de cada óvulo vegetal se encontra uma célula especializada, a oosfera, que é o gameta feminino propriamente dito.
Diagramas florais
O número dos tipos de peças florais estudadas é variável de flor para flor e pode ser representado esquematicamente por um diagrama. Cada tipo pode ser representado por 3, 4 ou 5 peças ou múltiplos desses números. Na flor do hibisco, por exemplo, uma planta comum em jardins, há 5 sépalas, 5 pétalas, um número múltiplo de 5 estames e um pistilo cujo ovário é dividido em 5 lojas.
Inflorescências
Em algumas plantas muitas flores se agrupam em um mesmo ramo, formando conjuntos denominados inflorescências.
 Flor do brócolis
Flor do brócolis
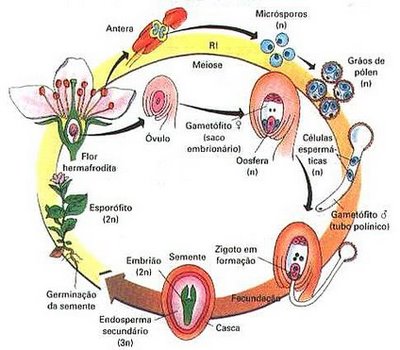
Formação dos frutos e das sementes
Para que servem as flores?
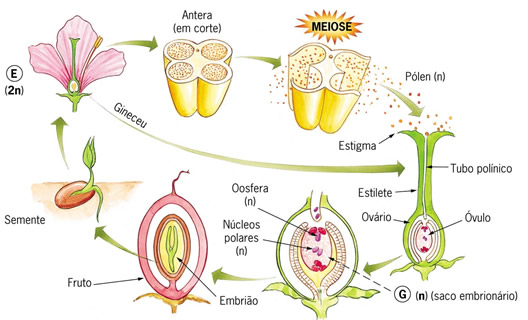
Após a polinização e a fecundação, a flor sofre uma modificação extraordinária. De todos os componentes que foram vistos anteriormente, acabam sobrando apenas o pedúnculo e o ovário. Todo o restante degenera. O ovário sofre uma grande modificação, se desenvolve e agora dizemos que virou fruto. Em seu interior os óvulos viram sementes.
Assim, a grande novidade das angiospermas, em termos de reprodução, é a presença dos frutos. Todos os componentes da flor que estudamos participa do processo reprodutivo que culminará na produção de sementes dentro do fruto. Em toda a angiosperma é assim, mas deve-se se lembrar que existe variações: há diferentes formatos de frutos e diferentes quantidades ou até mesmo nenhuma semente.
Quando a planta tem inflorescências para a reprodução, os frutos formados também ficarão reunidos e constituirão as infrutescências. É o caso do cacho de uvas, da amora, da jaca e da espiga de milho.
Como ocorre a formação dos frutos
Polinização e fecundação
Polinização é o transporte dos grãos de pólen das anteras, onde eles se formam, até o estigma, geralmente de uma outra flor. A polinização é o primeiro passo para a aproximação dos gametas femininos e masculinos, essencial para que a fecundação ocorra.
O transporte do pólen, até o estigma é feito por agentes polinizadores, que podem ser o vento, os insetos ou os pássaros.
Anemofilia
A polinização pelo vento é chamada de anemofilia (do grego anemos, vento). Há diversas adaptações que favorecem esse tipo de polinização. As flores de plantas anemófilas geralmente tem estigmas plumosos, que oferecem maior superfície para receber os grãos de pólen. Suas anteras geralmente possuem filetes longos e flexíveis que oscilam ao vento, o que facilita a dispersão do pólen. Além disso, as plantas anemófilas costumam produzir grande quantidade de grãos de pólen, o que aumenta as chances de polinização.
Entomofilia e ornitofilia
A polinização por insetos é chamada entomofilia (do grego entomos, inseto) e a polinização por aves, ornitofilia (do grego ornithos, aves). As flores polinizadas por animais geralmente possuem características que atraem os polinizadores, tais como corola vistosa, glândulas odoríferas e produtoras de substâncias açucaradas (néctar). Existem até mesmo flores que produzem dois tipos de estames, um com grãos de pólen férteis mas pouco atraentes e outro com pólen atraente e comestível. O animal à procura do pólen comestível, se impregna com o pólen fértil, transportando-o de uma flor para a outra.
Fecundação
Um grão de pólen, ao atingir o estigma de uma flor de mesma espécie, é estimulado a se desenvolver por substâncias indutoras presentes no estigma. O pólen forma um longo tubo, o tubo polínico, que cresce pistilo adentro até atingir o óvulo. Este possui um pequeno orifício nos tegumentos, denominado micrópila, por onde o tubo polínico penetra. Pelo interior do tubo polínico deslocam-se duas células haplóides, os núcleos espermáticos, que são os gametas masculinos.
No interior do óvulo há uma célula haplóide especial, a oosfera, que corresponde ao gameta feminino. A oosfera situa-se em posição estratégica dentro do óvulo, bem junto a pequena abertura denominada mocrópila. O tubo polínico atinge exatamente a micrópila ovular e um dos dois núcleos espermáticos do pólen fecunda a oosfera, originado o zigoto. Este dará origem ao embrião.
O outro núcleo espermático se une a dois núcleos polares presentes no interior do óvulo, originando um tecido triplóide, o endosperma, que nutrirá o embrião.
O óvulo fecundado se transforma na semente, que contém um pequeno embrião em repouse em seu interior.
Veja com mais detalhes!
Frutos e sementes
Os frutos surgem do desenvolvimento dos ovários, geralmente após a fecundação dos óvulos. Em geral, a transformação do ovário em fruta é induzida por hormônios liberados pelos embriões em desenvolvimento. Existem casos, porém, em que ocorre a formação de frutos sem que tenha havido polinização.
Partes do fruto
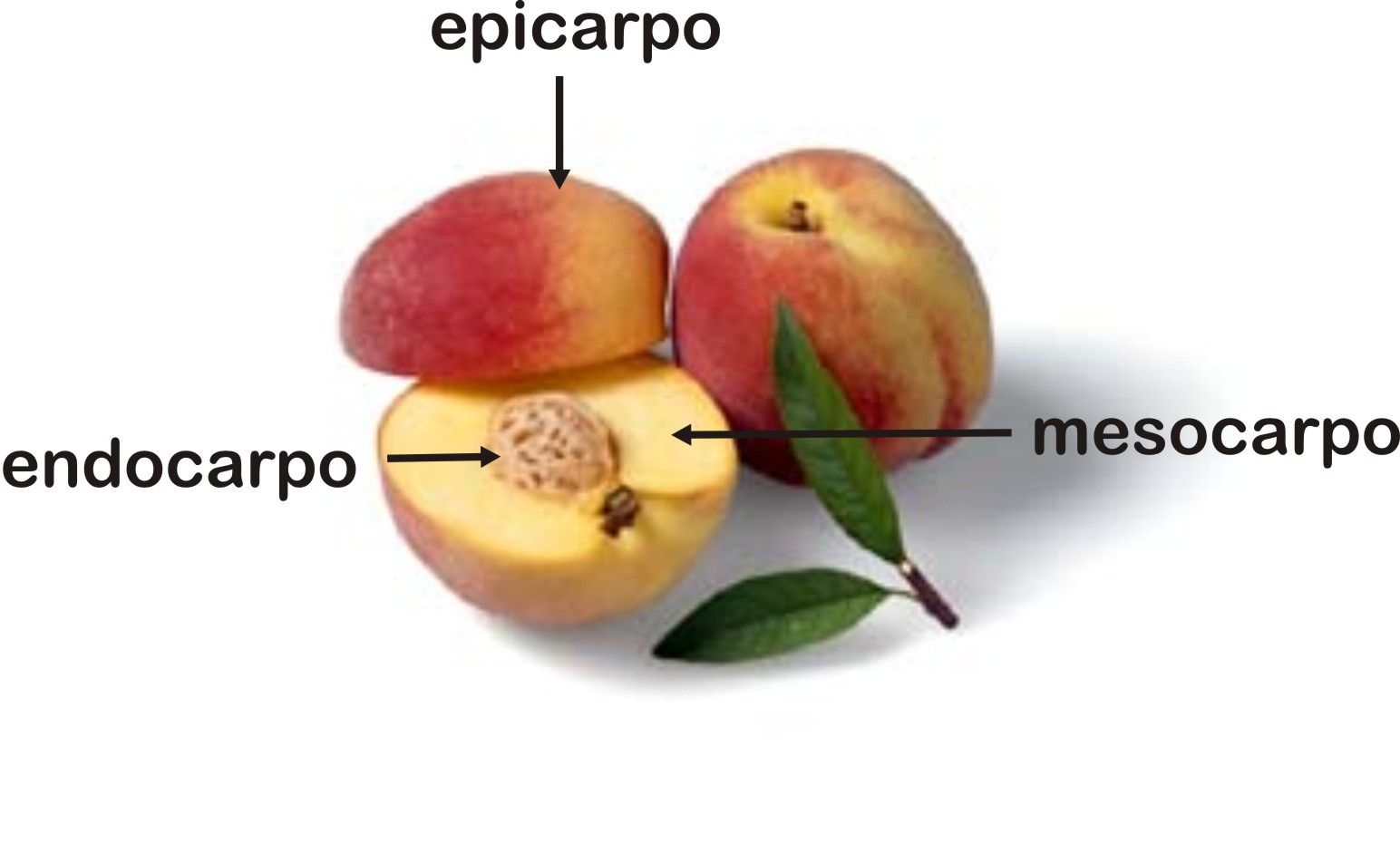
Um fruto é constituído por duas partes principais: o pericarpo, resultante do desenvolvimento das paredes do ovário, e as sementes, resultantes do desenvolvimento dos óvulos fecundados.
O pericarpo compõe-se de três camadas: epicarpo (camada mais externa), mesocarpo (camada intermediária) e endocarpo (camada mais interna). Em geral o mesocarpo é a parte do fruto que mais se desenvolve, sintetizando e acumulando substâncias nutritivas, principalmente açucares.
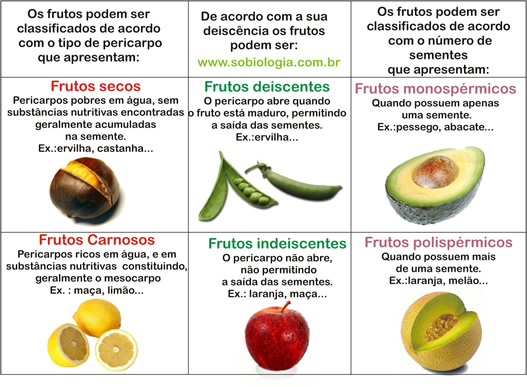
Classificação dos frutos
Diversas características são utilizadas para se classificar os frutos, entre elas o tipo de pericarpo, se o fruto abre-se ou não espontaneamente para liberar as sementes, etc.
Frutos que apresentam pericarpo suculento são denominados carnosos e podem ser do tipo baga, quando se originam de ovários uni ou multicarpelares com sementes livres (ex.: tomate, abóbora, uma e laranja), ou do tipo drupa, quando se originam de ovários unicarpelares, com sementes aderidas ao endocarpo duro (ex.: azeitona, pêssego, ameixa e amêndoa).
Frutos que apresentam endocarpo não suculento são chamados de secos e podem ser deiscentes, quando se abrem ao amadurecer, liberando suas sementes, ou indeiscentes, quando não se abrem ao se tornar maduros.
A diferença de fruta e fruto
O que se conhece popularmente por “frutas” não tem significado botânico. Fruta é aquilo que tem sabor agradável, às vezes azedo, às vezes doce. É o caso da laranja, pêssego, caju, banana, pêra, maça, morango, amora. Note que nem toda fruta é fruto verdadeiro.
Já o tomate, a berinjela, o jiló e a abobrinha, entre outros, são frutos verdadeiros, mas não são frutas...
Pseudofrutos e frutos partenocárpicos
Nos
pseudofrutos a porção comestível não corresponde ao ovário desenvolvido. No caju, ocorre hipertrofia do pedúnculo floral. Na maça, na pêra e no morango, é o receptáculo floral que se desenvolve.
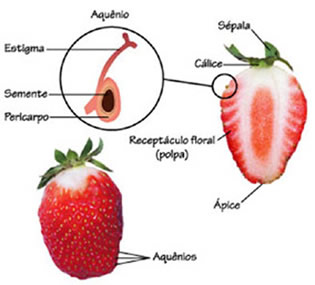
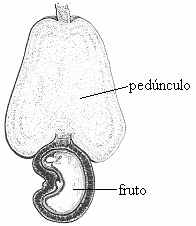
Assim, ao comer a polpa de um abacate ou de uma manga você está se alimentando do fruto verdadeiro. No entanto, ao saborear um caju ou uma maça, você está mastigando o pseudofruto.
No caso da banana e da laranja de umbigo (baiana), o fruto é partenocárpico, corresponde ao ovário desenvolvido sem fecundação, logo, sem sementes.
Origem e estrutura da semente
A semente é o óvulo modificado e desenvolvido. Toda a semente possui um envoltório, mais ou menos rígido, um embrião inativo da futura planta e um material de reserva alimentar chamado endosperma ou albúmen.
Em condições ambientais favoráveis, principalmente de umidade, ocorre a hidratação da semente e pode ser iniciada a germinação.
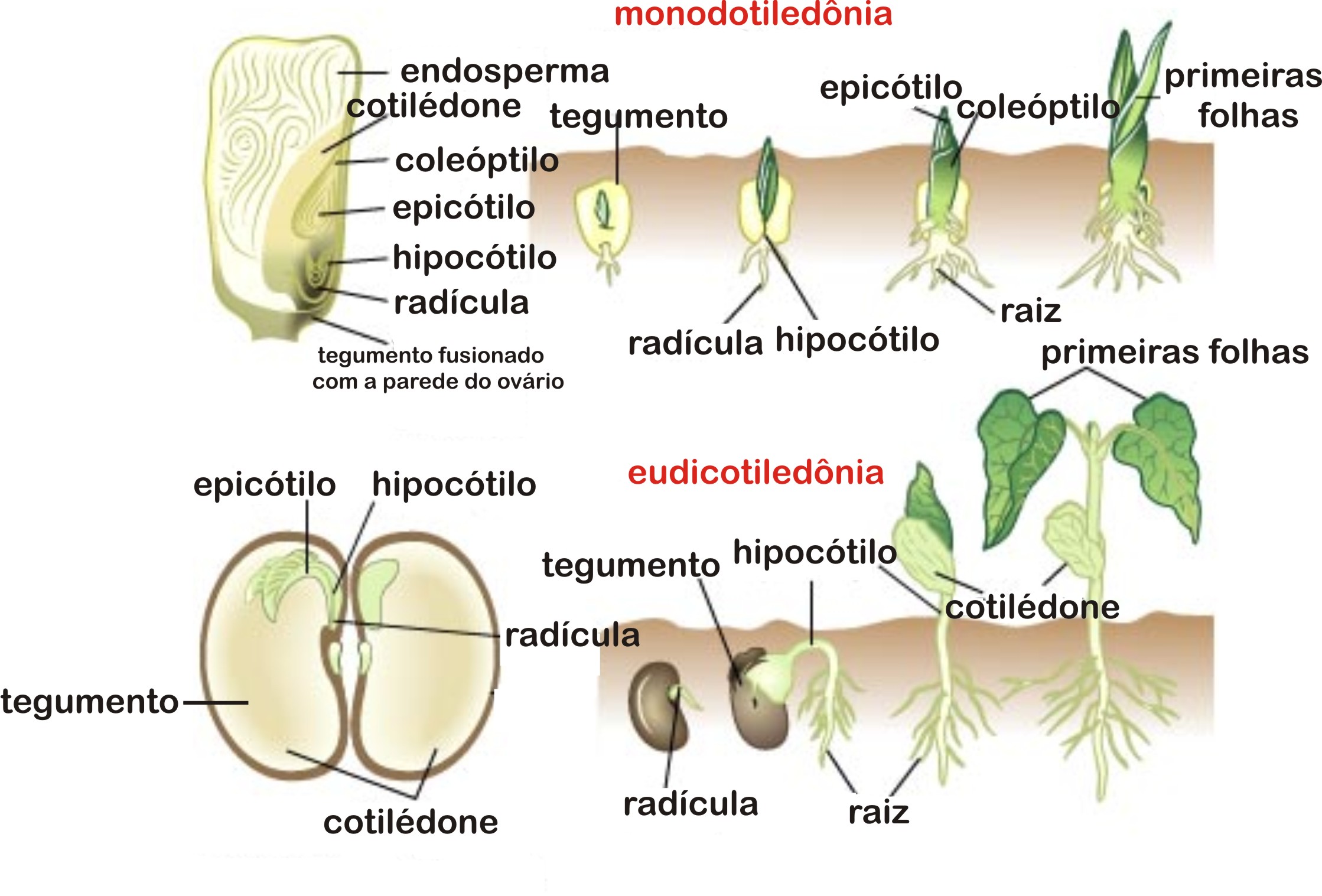
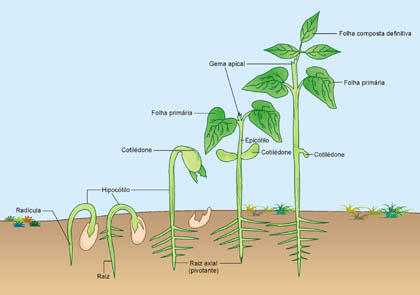
Os cotilédones
Todo o embrião contido em uma semente de angiosperma é um eixo formado por duas extremidades:
- A radícula, que é a primeira estrutura a emergir, quando o embrião germina;e
- O caulículo, responsável pela formação das primeiras folhas embrionárias.
Uma “folha” embrionária merece especial atenção. É o cotilédone. Algumas angiospermas possuem dois cotilédones, outras possuem apenas um. Plantas que possuem dois cotilédones, são chamadas de eudicotiledôneas e plantas que possuem um cotilédone sã chamadas de monocotiledôneas. Os cotilédones inserem-se no caulículo, que dará origem ao caule.
A célula vegetal
A compreensão da anatomia e da fisiologia das plantas depende, fundamentalmente, do conhecimento sobre a organização e o funcionamento de suas células.
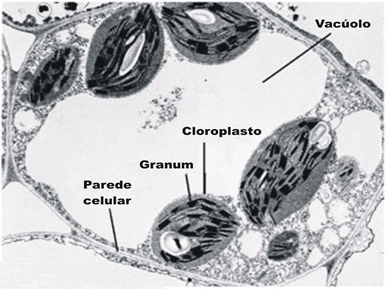
As células das plantas vegetais apresentam pelo menos duas características que permitem distinguilas claramente das células animais: possuem um envoltório externo rígido, a parede celular, e um orgânulo citoplasmático responsável pela fotossíntese, o plasto. Além disso, quando adultas, a maioria das células vegetais possui uma grande bolsa membranosa na região central do citoplasma, o vacúolo central, que acumula uma substância aquosa de sais e açúcares.
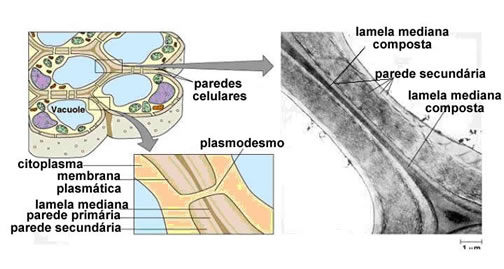
Parede da célula vegetal
A parede celular começa a se formar ainda na telófase da mitose que dá origem à célula vegetal. Bolsas membranosas oriundas do aparelho de Golgi, repletas de substâncias gelatinosas denominadas pectinas, acumulam-se na região central da célula em divisão e se fundem, originando uma placa chamadafragmoplasto.
Enquanto a telófase avança, o fragmoplasto vai crescendo pela fusão de bolsas de pectina em suas bordas. Durante esse crescimento centrífugo (isto é, do centro para fora), forma-se poros no fragmoplasto, por onde passa fios de hialoplasma, que põe em comunicação os conteúdos das futuras células vizinhas. Essas pontes hiloplasmáticas são os plasmosdesmos (do grego plasmos, líquido, relativo ao citoplasma, e desmos, ponte, união).
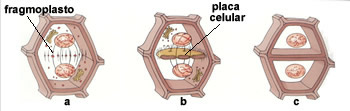
O fragmoplasto atua como uma espécie de “forma” para a construção das paredes celulósicas. Cada célula irmã-secreta celulose sobre o fragmoplasto e vai construindo, de seu lado, uma parede celulósica própria. A camada de pectinas, que foi a primeira separação entre as células-irmãs, atua agora como um cimento intercelular, passando a se chamar lamela média.
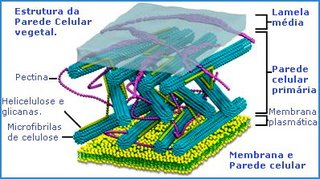
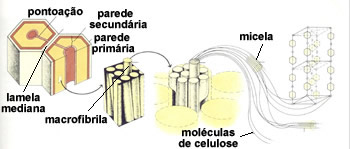
Estrutura da parede celular vegetal
A parede da célula vegetal é constituída por longas e resistentes microfibrilas de celulose. Uma microfibrila reúne entre sessenta e setenta moléculas de celulose, cada qual, constituída, por sua vez, por quinhentas moléculas de glicose encadeadas linearmente. As microfibrilas de celulose mantêm-se unidas por uma matriz formada por glicoproteínas (proteínas ligadas á açucares) e por dois polissacarídeos,hemicelulose e pectina. Esta estruturação molecular lembra o concreto armado, onde longas e resistentes varetas de ferro, correspondentes as microfibrilas celulósicas, ficam mergulhadas em uma argamassa de cimento e pedras, correspondente à matriz de glicoproteínas, hemicelulose e pectina.
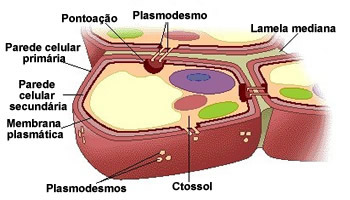
Paredes primárias e secundárias
A parede celulósica secretada logo após a divisão celular é a parede primária. Essa parede é elástica e acompanha o crescimento celular. Depois que a célula atingiu o seu tamanho e forma definitivos, ela secreta uma nova parede internamente à parede primária. Essa é a parede secundária.
As especializações das células das plantas estão sempre associadas à estrutura das paredes celulares. Nos diferentes tecidos vegetais as células têm paredes diferentes de diferentes espessuras, organização e composição química, que determinam não só a forma como também as funções das células.
Conteúdo celular - principais organelas
Vacúolo
Delimitado por uma membrana denominada tonoplasto. Contém água, açúcares, proteínas; pode-se encontrar ainda compostos fenólicos, pigmentos como betalaínas, antocianinas cristais de oxalato de cálcio (drusas, estilóides, cristais prismáticos, rafídios, etc.). Muitas das substâncias estão dissolvidas, constituindo o suco celular, cujo pH é geralmente ácido, pela atividade de uma bomba de próton no tonoplasto. Em células especializadas pode ocorrer um único vacúolo, originado a partir da união de pequenos vacúolos de uma antiga célula meristemática (célula-tronco); em células parenquimáticas o vacúolo chega a ocupar 90% do espaço celular.
Funções: Ativo em processos metabólicos, como:
- armazenamento de substâncias (vacúolos pequenos - acúmulo de proteínas, íons e outros metabólitos). Um exemplo são os microvacúolos do endosperma da semente de mamona (Ricinus communis), que contêm grãos de aleurona;
- processo lisossômico (através de enzimas digestivas, existentes principalmente nos vacúolos centrais e bem desenvolvidos, cujo tonoplasto sofre invaginações para englobar material citoplasmático contendo organelas (a autofagia ocorre em células jovens ou durante a senescência). Se originam a partir do sistema de membranas do complexo golgiense. Seu tamanho aumenta à medida que o tonoplasto incorpora vesículas derivadas do complexo de Golgi.
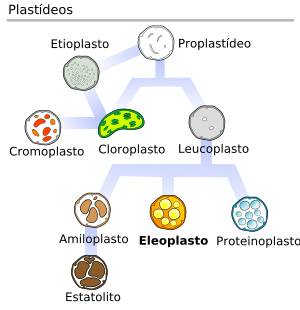
Plastos
Organelas formadas por um envelope de duas membranas unitárias contendo internamente uma matriz ou estroma, onde se situa um sistema de membranas saculiformes achatadas, os tilacóides. Originam-se dos plastídios e contêm DNA e ribossomos.
São divididos em três grandes grupos:
- cloroplasto;
- cromoplasto e
- leucoplasto; estes, por sua vez, originam-se de estruturas muito pequenas, os proplastídios (que normalmente já ocorrem na oosfera, no saco embrionário e nos sistemas meristemáticos). Quando os proplastídios se desenvolvem na ausência de luz, apresentam um sistema especial, derivado da membrana interna, originando tubos que se fundem e formam o corpo prolamelar. Esses plastos são chamados estioplastos.
Cloroplastos: Seu genoma codifica algumas proteínas específicas dessas organelas; contêm clorofila e estão associados à fase luminosa da fotossíntese, sendo mais diferenciados nas folhas. Seu sistema de tilacóides é formado por pilhas de membranas em forma de discos, chamado de granus; é nesse sistema que se encontra a clorofila. Na matriz ocorrem as reações de fixação de gás carbônico para a produção de carboidratos, além de aminoácidos, ácidos graxos e orgânicos. Pode haver formação de amido e lipídios, estes últimos em forma de glóbulos (plastoglóbulos).
Cromoplastos: Portam pigmentos carotenóides (geralmente amarelos, alaranjados ou avermelhados); são encontrados em estruturas coloridas como pétalas, frutos e algumas raízes. Surgem a partir dos cloroplastos.
Leucoplastos: Sem pigmentos; podem armazenar várias substâncias:
- amiloplastos: armazenam amido. Ex.: em tubérculos de batatinha inglesa (Solanum tuberosum).
- proteinoplastos: armazenam proteínas.
- elaioplastos: armazenam lipídios. Ex.: abacate (Persea americana).
Tecidos vegetais
Um violento temporal, uma seca prolongada, um animal herbívoro ou qualquer outro agente agressivo do meio, têm que ser enfrentados pela planta imóvel, ao contrário de um animal, que pode se refugiar em lugar seguro até que as condições ambientais se normalizem.
Os tecidos protetores, ou de revestimento, de uma traqueófita são a epiderme e o súber. A eficiência deles pode garantir a proteção da planta contra diversos agentes agressivos do meio.
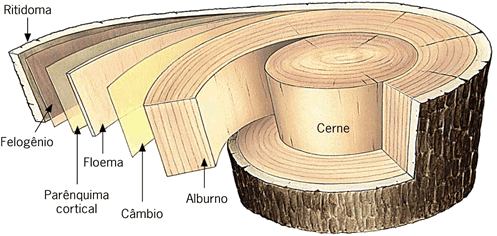
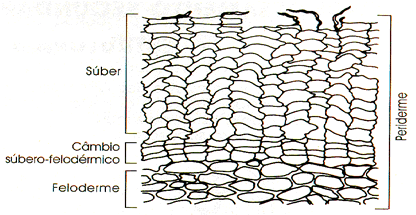
O súber
É um tecido de revestimento existente em raízes e troncos – portanto em plantas arborescentes adultas, espesso, formado por várias camadas de células mortas. A morte celular, nesse caso é devida a impregnação de grossas camadas de suberina (um material lipídico) nas paredes da célula que fica, assim, oca. Como armazena ar, o súber funciona como um excelente isolante térmico, além de exercer, é claro, um eficiente papel protetor.
 Ritidoma
Ritidoma
O tronco de uma árvore periodicamente cresce em espessura. Esse crescimento força a ruptura do súber que racha em muitos pontos e acaba se destacando, juntamente com outros tecidos. Antes, porém, a árvore elabora novo súber que substituirá o que vai cair. A este material periodicamente destacado dá-se o nome de ritidoma.
A epiderme
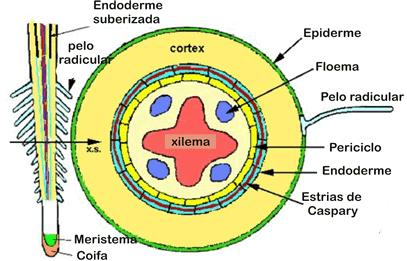
A epiderme das plantas vasculares é um tecido formado, de modo geral, por uma única camada de células de formato irregular, achatadas, vivas e aclorofiladas. É um tecido de revestimento típico de órgãos jovens (raiz, caule e folhas). A epiderme de uma raiz mostra uma camada cilíndrica de revestimento, com uma zona pilífera, cujos pelos nada mais são do que extensões de uma célula epidérmica.
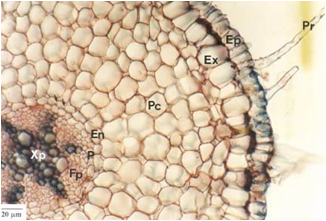 Corte transversal da raiz primária de Mandevilla velutina. Ep = epiderme; Pr = pêlos radicular; Ex = exoderme; Pc = parênquima cortical; En = endoderme; P = periciclo; Xp = xilema primária; Fp = floema primário.
Corte transversal da raiz primária de Mandevilla velutina. Ep = epiderme; Pr = pêlos radicular; Ex = exoderme; Pc = parênquima cortical; En = endoderme; P = periciclo; Xp = xilema primária; Fp = floema primário.
Caules jovens também são revestidos por uma fina epiderme não-dotada, porém, de pelos.
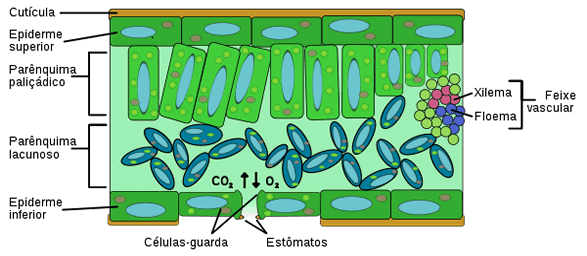
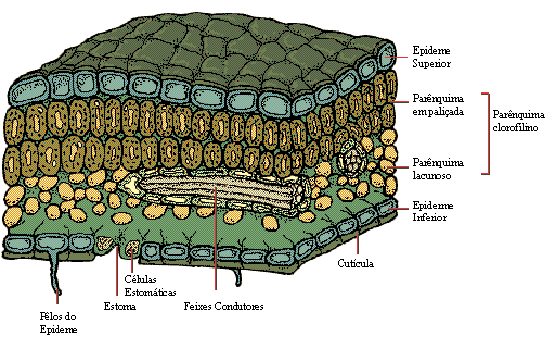
É na folha que a epiderme possui notáveis especializações: sendo um órgão de face dupla, possui duas epidermes, a superior e a inferior.
As células epidérmicas secretam para o exterior substâncias impermeabilizantes, que formam uma película de revestimento denominada cutícula. O principal componente da cutícula é a cutina, um polímero feito de moléculas de ácidos graxos. Além de evitar a perda de água, a cutícula protege a planta contra infecções e traumas mecânicos.
Os anexos da epiderme
Diferenciam-se na epiderme estruturas como estômatos, tricomas, hidatódios e acúleos.
Estômatos
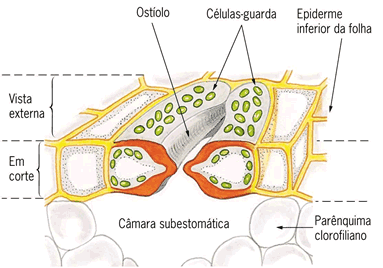
Sem dúvida, os estômatos são os anexos mais importantes relacionados com a troca de gases e água entre as folhas e o meio. As células estomáticas são as únicas na epiderme que possuem clorofila. Um estômato visto de cima, assemelha-se a dois feijões dispostos com as concavidades frente a frente: são as duas células estomáticas ou células-guarda, que possuem parede celular mais espessa na face côncava e cuja disposição deixa entre elas um espaço denominado fenda estomática ou ostíolo.
Ao lado de cada célula-guarda há uma anexa, que não tem cloroplastos – é uma célula epidérmica comum. Em corte transversal, verifica-se que a fenda estomática dá acesso a um espaço, a câmara estomática, intercomunicante com os espaços aéreos do parênquima foliar de preenchimento.
Atenção! A troca de gases entre a planta e o meio ocorre através dos estômatos da epiderme e de uma estrutura chamada lenticelas presentes no súber.
As lenticelas são pequenas aberturas que facilitam o ingresso e a saída de gases nas raízes e caules suberificados.
|
Tricomas
Os tricomas são geralmente estruturas especializadas contra a perda de água por excesso de transpiração, ocorrendo em planta de clima quente. Podem ser, no entanto, secretores, produzindo secreções oleosas, digestivas ou urticantes. As plantas carnívoras possuem tricomas “digestivos” e a urtiga, planta que provoca irritação da pele, possui tricomas urticantes.
Acúleos
Os acúleos, estruturas pontiagudas com função de proteção da planta contra predadores, são frequentemente confundido com espinhos, que são folhas ou ramos modificados. Os acúleos são fáceis de destacar e são provenientes da epiderme. Podem ser encontrados nas roseiras.
Hidatódios
Hidatódios são estômatos modificados, especializados em eliminar excessos líquidos da planta. Os hidatódios geralmente presentes nas bordas das folhas, onde, pela manhã, é possível observar as gotas de líquido que eles eliminam, fenômeno conhecido como gutação.
A sustentação das traqueófitas
O porte das traqueófitas só foi possível por adaptações que tornaram possível a sustentação do organismo vivo e a disponibilidade e transporte de água para todas as células.
A sustentação de uma traqueófita é devida à existência de tecidos especializados para essa função: ocolênquima e o esclerênquima.
O Colênquima
As células do colênquima são alongadas, irregulares e encontram-se dispostas em forma de feixes. Quando cortadas transversalmente, têm aspecto variado. São vivas, nucleadas, e a parede apresenta reforços de celulose, mais intensos nos cantos internos da célula, conferindo certa resistência ao esmagamento lateral. O colênquima é um tecido flexível, localizado mais externamente no corpo do vegetal e encontrado em estruturas jovens como pecíolo de folhas, extremidade do caule, raízes, frutos e flores.
O Esclerênquima
O esclerênquima é um tecido mais rígido que o colênquima, encontrado em diferentes locais do corpo de uma planta. As células do esclerênquima possuem um espessamento secundário nas paredes devido à impregnação de lignina. As células mais comuns do esclerênquima são as fibras e os esclerídeos, também chamados escleritos.
Estrutura interna das folhas
A folha é totalmente revestida pela epiderme, e seu interior, denominado mesófilo (do grego, mesos, meio e phylon, folha), é constituído por parênquima clorofiliano, tecidos condutores e tecidos de sustentação.
O parênquima clorofiliano foliar pode ser, em geral de dois tipos:
- palisádico - constituído por células prismáticas e justapostos como uma paliçada, e
- lacunoso -constituído por células de forma irregular, que deixam espaços ou lacunas entre si.
Pode haver parênquima paliçádico junto à epiderme de ambas as faces da folha, ou, como é mais comum, parênquima paliçádico junto a epiderme da face superior e lacunoso junto à inferior.
Nervuras foliares
Os tecidos condutores presentes na folha encontram-se agrupados em feixes libero-lenhosos, nos quais o xilema está voltado para a epiderme superior e o floema, para a epiderme inferior. Os feixes condutores mais grossos formam as nervuras foliares, visíveis a olho nu.
Os Tecidos Condutores de Água e de Nutrientes em Traqueófitas
Além das trocas gasosas, um dos maiores problemas de um vegetal terrestre relaciona-se à disponibilidade de água e sua perda, pois para à realização da fotossíntese é fundamental que se consiga, além do gás carbônico, a água. O problema de perda de água através das folhas é, em parte, minimizado pela presença de cutículas lipídicas, nas faces expostas das epidermes, que as impermeabilizam. Porém, isso dificulta as trocas gasosas.
A existência nas traqueófitas de aberturas epidérmicas reguláveis (os estômatos) que permitem as trocas gasosas e ao mesmo tempo ajudam a evitar perdas excessivas de vapor de água é um mecanismo adaptativo importante.
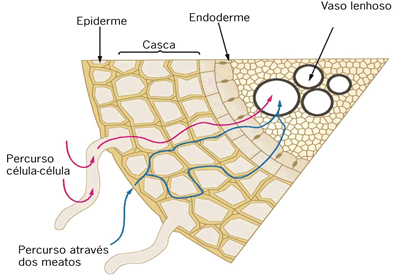
O transporte de água e nutrientes em uma traqueófita ocorre em parte por difusão de célula à célula e, na maior parte do trajeto, ocorre no interior de vasos condutores.
Inicialmente, ocorre a absorção de água e nutrientes minerais pela zona pilífera da raiz. Os diferentes tipos de íons são obtidos ativa ou passivamente e a água é absorvida por osmose.
Forma-se uma solução aquosa mineral, a seiva bruta ou seiva inorgânica. Essa solução caminha de célula a célula radicular até atingir os vasos do xilema (ou lenho) existentes no centro da raiz. A partir daí, o transporte dessa seiva ocorre integralmente dentro dos vasos lenhosos até as folhas. Lá chegando, os nutrientes e a água difundem-se até as células e são utilizados no processo da fotossíntese.
Os compostos orgânicos elaborados nas células do parênquima clorofiliano das folhas difundem-se para outro conjunto de vasos do tecido condutor chamado floema ou líber. No interior dos vasos liberianos, essa seiva orgânica ou seiva elaborada é conduzida até atingir as células do caule, de um fruto, de um broto em formação, de uma raiz etc., onde é utilizada ou armazenada.
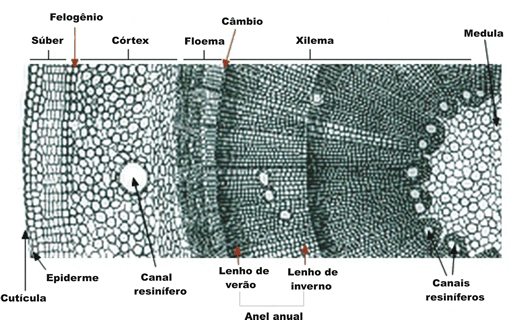
O xilema
Os vasos condutores de seiva inorgânica são formados por células mortas. A morte celular é devida à impregnação da célula por lignina, um composto aromático altamente impermeabilizante. A célula deixa de receber nutrientes e morre. Desfaz-se o conteúdo interno da célula, que acaba ficando oca e com as paredes duras já que a lignina possui, também, a propriedade de endurecer a parede celular. A deposição de lignina na parede não é uniforme. A célula, então, endurecida e oca, serve como elemento condutor. Existe, ainda, um parênquima (tecido vivo) interposto que separa grupos de células condutoras. Acredita-se que essas células parenquimáticas secretem diferentes tipos de substâncias que provavelmente auxiliam a preservação dos vasos mortos do xilema.
Existem dois tipos de células condutoras no xilema: traqueíde e elemento de vaso traqueário (ou xilemático ou, ainda, lenhoso).
 |
- Traqueídes são células extremamente finas, de pequeno comprimento (em média 4 mm) e diâmetro reduzido (da ordem de 2 mm). Quando funcionais, as traqueídes estão agrupadas em feixes e as extremidades de umas tocam as das outras. Na extremidade de cada traqueíde, assim como lateralmente, há uma série de pontuações ou poros(pequeníssimos orifícios) que permitem a passagem de seiva no sentido longitudinal e lateral.
- Menores que as traqueídes (em média de 1 a 3 mm), porém mais longos (até 300 mm), oselementos de vaso também possuem pontuações laterais que permitem a passagem da seiva. Sua principal característica é que em suas extremidades as paredes são perfuradas, isto é, não há parede divisória totalmente isolante entre uma e outra célula. O vaso formado pela reunião de diversos elementos de vaso é conhecido como traquéia.
O nome traquéia para o vaso condutor é derivado da semelhança que os reforços de lignina do vaso apresentam com os reforços de cartilagem da traquéia humana e os de quitina dos insetos.
|
A condução da seiva inorgânica
Vimos que as raízes absorvem água do solo através da região dos pelos absorventes ou zona pilífera. Desta, a água atravessa as células do córtex, endoderme e periciclo da raiz. Na endoderme o fluxo da água pode ser facilitado ela existência das chamadas células de passagem. A água atinge os vasos do xilema e, a partir desses vasos, atinge a folha. Na folha, ou ela é usada na fotossíntese ou é liberada na transpiração.
Atribui-se a condução da seiva inorgânica (ou bruta) a alguns mecanismos: pressão da raiz, sucção exercida pelas folhas e capilaridade.
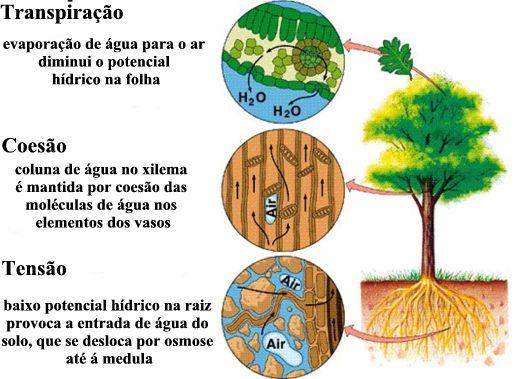
- A pressão da raiz – O movimento da água através da raiz é considerado como resultante de um mecanismo osmótico. A água que está no solo entra na célula do pêlo radicular, cuja concentração é maior que a da solução do solo. A célula radicular é menos concentrada que a célula cortical. Esta, por sua vez, é menos concentrada que a célula endodérmica e, assim por diante, até chegar ao vaso do xilema, cuja solução aquosa é mais concentrada de todas nesse nível. Assim, é como se a água fosse osmoticamente bombeada, até atingir os vasos do xilema.
- A sucção exercida pelas folhas – A hipótese mais aceita, atualmente, para o deslocamento da seiva do xilema é baseada na “sucção” de água que a copa exerce. Esta “sucção” está relacionada com os processos de transpiração e fotossíntese que ocorrem nas folhas. Para que essa “aspiração” seja eficiente, dois pré-requisitos são fundamentais: inexistência de ar nos vasos de xilema e uma força de coesão entre as moléculas de água. A coesão entre as moléculas de água faz com que elas permaneçam unidas umas às outras e suportem forças extraordinárias, como o próprio peso da coluna líquida no interior dos vasos, que poderiam levá-las a separar-se. A existência de ar nos vasos do xilema romperia essa união e levaria à formação de bolhas que impediriam a ascensão da seiva lenhosa. As paredes dos vasos lenhosos igualmente atraem as moléculas de água e essa adesão, juntamente com a coesão, são fatores fundamentais na manutenção de uma nova coluna contínua de água no interior do vaso.
- A transpiração e a fotossíntese removem constantemente água da planta. Essa extração gera uma tensão entre as moléculas de água já que a coesão entre elas impede que se separem. A parede do vaso também é tracionada devido à adesão existente entre ela e as moléculas de água. Para que se mantenha a continuidade da coluna líquida, a reposição das moléculas de água retiradas da copa deve ser feita pela raiz, que, assim, abastece constantemente o xilema.
- O efeito da capilaridade na condução da seiva – Os vasos lenhosos são muito delgados, possuem diâmetro capilar. Assim, a ascensão do xilema ocorre, em parte, por capilaridade. No entanto, por esse mecanismo, a água atinge alturas bem inferiores a 1 metro e, isoladamente, esse fato é insuficiente para explicar a subida da seiva inorgânica.
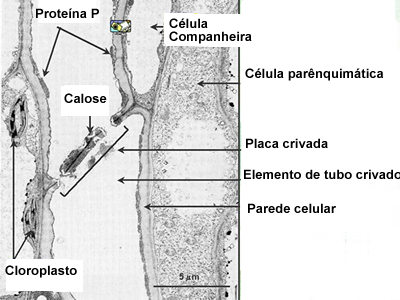
O floema
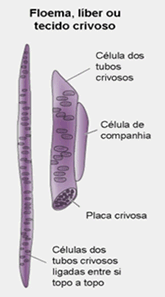 | |
Os vasos do floema (também chamado de líber) são formados porcélulas vivas, cuja parede possui apenas a membrana esquelética celulósica típica das células vegetais e uma fina membrana plasmática. São células altamente especializadas e que perdem o núcleo no decorrer do processo de diferenciação. O seu interior é ocupado pela seiva elaborada (ou seiva orgânica) e por muitas fibras de proteínas, típicas do floema. A passagem da seiva orgânica de célula a célula é facilitada pela existência de placas crivadas nas paredes terminais das células que se tocam. Através dos crivos, flui a seiva elaborada de uma célula para outra, juntamente com finos filamentos citoplasmáticos,os plasmodesmos.
Os orifícios das placas crivadas são revestidos por calose. Polissacarídeo que obstrui os crivos quando, em alguns vegetais, periodicamente, os vasos crivados ficam sem função. Ao retornarem à atividade, esse calo é desfeito.
Lateralmente aos tubos crivados, existem algumas células delgadas, nucleadas, chamadas de companheiras, cujo núcleo passa a dirigir também a vida das células condutoras.
|
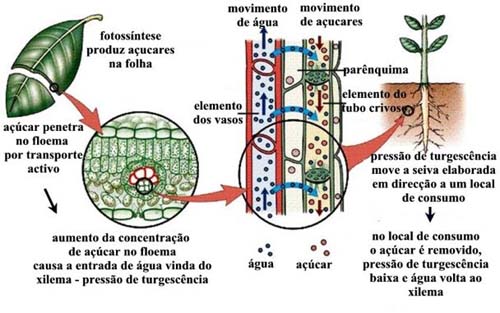
A condução da seiva elaborada
A seiva orgânica, elaborada no parênquima das folhas, é lançada nos tubos crivados do floema e conduzida a todas as partes da planta que não são auto-suficientes. O transporte é orientado principalmente para a raiz, podendo haver algum movimento em direção ao ápice do caule e folhas em desenvolvimento. De modo geral, os materiais orgânicos são translocados para órgãos consumidores e de reserva, podendo haver inversão do movimento (isto é, dos órgãos de reserva para regiões em crescimento), quando necessário.
A hipótese de Münch
A hipótese mais aceita atualmente para a condução da seiva elaborada é a que foi formulada por Münch e se baseia na movimentação de toda a solução do floema, incluindo água e solutos. É a hipótese doarrastamento mecânico da solução, também chamada de hipótese do fluxo em massa da solução. Por essa hipótese, o transporte de compostos orgânicos seria devido a um deslocamento rápido de moléculas de água que arrastariam, no seu movimento, as moléculas em solução.
A compreensão dessa hipótese fica mais fácil acompanhando-se o modelo sugerido por Münch para a sua explicação.
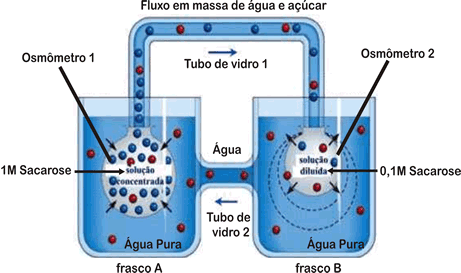
Observando a figura, conclui-se que haverá ingresso de água por osmose, do frasco A para o osmômetro 1, e do frasco B para o osmômetro 2. No entanto, como a solução do osmômetro 1 é mais concentrada, a velocidade de passagem de água do frasco A para o osmômetro 1 é maior. Assim, a água tenderá a se dirigir para o tubo de vidro 1 com velocidade, arrastando moléculas de açúcar. Como o osmômetro 2 passa a receber mais água, esta passa para o frasco B. Do frasco B, a água passa para o tubo de vidro 2, em direção ao frasco A. Podemos fazer a correspondência entre o modelo anterior e uma planta:
- Tubo de vidro 1 corresponde ao floema e o tubo de vidro 2 ao xilema;
- Osmômetro 1 corresponde a uma célula do parênquima foliar e o osmômetro 2, a uma célula da raiz;
- Frasco A representa a folha, enquanto o frasco B representa a raiz;
- As células do parênquima foliar realizam fotossíntese e produzem glicose. A concentração dessas células aumenta, o que faz com que absorvam água do xilema das nervuras. O excesso de água absorvida é deslocado para o floema, arrastando moléculas de açúcar em direção aos centros consumidores ou de reserva.
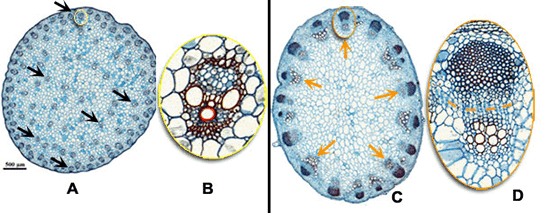
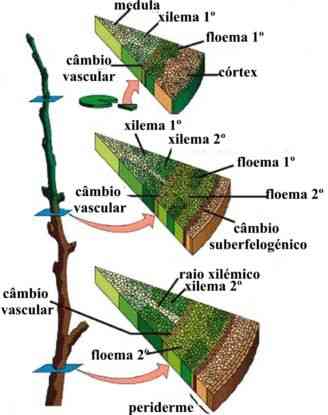
Organização dos Tecidos nas Raízes e nos Caules
Raízes e caules jovens, cortados transversalmente, mostram que são formados por uma reunião de tecidos. A disposição desses tecidos é específica em cada órgão e constitui uma estrutura interna primária típica de cada um deles. Uma estrutura secundária, mais complexa, pode ser vista quando ocorre um aumento no diâmetro do caule e da raiz.
Estrutura primária da raiz
Se acompanhássemos uma célula meristemática que terminou de surgir por mitose na extremidade de uma raiz, veríamos que ela vai se alongando, ao mesmo tempo que vai se distanciando da extremidade em decorrência do surgimento de novas células. A maior taxa de crescimento em extensão de uma raiz, ocorrerá, portanto, na região situada pouco acima da região meristemática, denominada de zona de distensão.
Após crescerem as células iniciam a sua diferenciação. Na região mais interna, por exemplo, terá início a diferenciação dos tecidos condutores, enquanto na região mais externa diferenciam-se parênquimas e tecidos de revestimento.
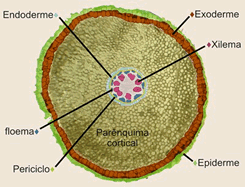
Córtex
A região mais periférica da raiz jovem diferenciam-se em epiderme, tecido formado por uma única camada de células achatadas e justapostas. Na região abaixo da epiderme, chamada córtex, diferencia-se o parênquima cortical, constituído por várias camadas de células relativamente pouco especializadas.
Cilindro central
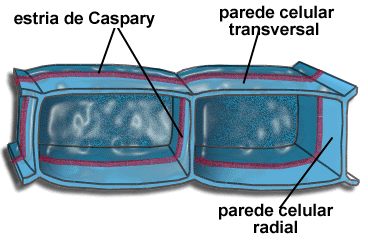
A parte interna da raiz é o cilindro central, composto principalmente por elementos condutores(protoxilema e protofloema), fibras e parênquima. O cilindro central é delimitado pela endoderme, uma camada de células bem ajustadas e dotadas de reforços especiais nas paredes, as estrias de Caspary. Essas estrias são como cintas de celulose que unem firmemente as células vizinhas, vedando completamente os espaços entre elas. Assim, para penetrar no cilindro central, toda e qualquer substância tem que atravessar diretamente as células endodérmicas, uma vez que as estrias de caspary fecham os interstícios intercelulares.
Logo abaixo da endoderme situa-se uma camada de células de paredes finas chamada periciclo, que delimita o cilindro central, onde se localizam o xilema e o floema. A maneira como os tecidos condutores se dispõem no cilindro central é um dos critérios para distinguir dicotiledôneas de monocotiledôneas.
Raízes de mono e dicotiledôneas
Na maioria das plantas dicotiledôneas o xilema se concentra na região mias interna do cilindro central. Quando se observa um corte transversal à raiz, vê-se que o protoxilema ocupa uma área em forma de cruz ou estrela, cujas pontas encostam no periciclo. O protofloema encontra-se nos vértices formados pelos “braços” da cruz. Entre o protoxilema e o protofloema há um meristema primário chamado procâmbio. Os demais espaços dentro do cilindro central são preenchidos por parênquima.
Nas plantas monocotiledôneas, o centro da raiz é ocupado por uma medula constituída por parênquima medular e os vasos lenhosos e liberianos dispõem-se ao redor.
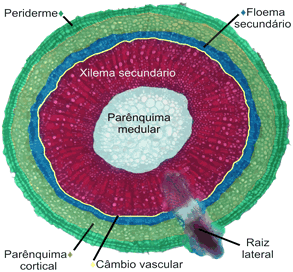
Estrutura secundária
O crescimento em espessura da raiz pode ser chamado de crescimento secundário, para distingui-lo do crescimento em extensão. Em linhas gerais, durante o crescimento secundário desenvolvem-se cilindros de células meristemáticas que permitem o surgimento de novos tecidos radiculares.
Os dois tecidos meristemáticos envolvidos no crescimento secundário da raiz são o câmbio vascular, que permite o crescimento do cilindro central, e o câmbio suberógeno ou felogênio, que permite o crescimento da periderme (casca).
Câmbio vascular
O câmbio vascular (do latim vasculum, vaso) é assim chamado porque origina novos vasos condutores durante o crescimento secundário da raiz. O câmbio vascular forma-se a partir do procâmbio e do periciclo, que se conjugam e delimitam uma área interna do cilindro central, onde só há xilema. Ao se multiplicar ativamente, as células do câmbio vascular originam vasos xilemáticos para a região mais interna e vasos floemáticos para a região mais externa. Aos poucos a área delimitada pelo câmbio vai tornando-se cada vez mais cilíndrica.
O cambio vascular da raiz é um meristema de origem mista, primária e secundária. Isso porque tem origem tanto no procâmbio, um meristema primário, quanto do periciclo, um tecido já diferenciado que sobre desdiferenciação.
Câmbio suberógeno ou felogênio
O câmbio suberógeno, também chamado de felogênio (do grego phellos, cortiça, e genos, que gera), é um cilindro de células meristemáticas localizado na região cortical da raiz, sob a epiderme. O felogênio é um meristema secundário, uma vez que tem origem por desdiferenciação de células do parênquima cortical.
Como vimos, a atividade do felogênio produz feloderme e súber, este último um tecido morto que protege externamente raízes e caules com crescimento secundário.
Estrutura interna do caule
Como na raiz, a parte mais jovem de um caule é a que se localiza junto à extremidade, onde ocorre a multiplicação das células do meristema apical, que permite o crescimento em extensão.
Logo abaixo da zona meristemática apical as células iniciam o processo de diferenciação celular, que leva ao aparecimento dos diversos tecidos que compõem o caule.
Origem das gemas axilares
Nem todas as células produzidas pelo meristema apical sofrem diferenciação. À medida que o caule cresce, permanecem grupos de células meristemáticas sob a epiderme, pouco acima do ponto de inserção das folhas. Esses grupos de células formam, nas axilas das folhas, protuberâncias chamadas gemas axilares ou laterais.
Estrutura primária
Feixes líbero-lenhosos
Caules que não cresceram, em espessura apresentam estrutura primária, caracterizada pela presença de feixes líbero-lenhosos localizados entre as células do parênquima que preenchem seu interior.
Cada feixe libero-lenhoso possui elementos do líber (floema) voltados para fora e elementos do lenho (xilema) voltados para dentro.
Nas plantas monocotiledôneas, que geralmente na apresentam crescimento secundário, os feixes condutores são distribuídos de maneira difusa no interior do caule. Já nas dicotiledôneas os feixes líbero-lenhosos distribuem-se regularmente, formando um cilindro.
Câmbio fascicular
Nos feixes libero-lenhosos das dicotiledôneas, o floema está voltado para o exterior do caule e o xilema para o interior. Entre o floema e o xilema de um feixe há um tecido meristemático: o câmbio vascular e o câmbio suberógeno ou felogênio.
Câmbio vascular
O Câmbio vascular do caule forma-se a partir do câmbio fascicular e do câmbio interfascicular, este último um tecido meristemátco secundário, resultante da dedisferenciação de células parenquimáticas localizadas entre os feixes libero-lenhosos. O câmbio fascicular passa a delimitar, assim, uma área interna do caule onde só há xilema e parênquima. Como na raiz, as células do câmbio vascular originam vasos xilemáticos para a região mais interna e vasos floemáticos para a região mais externa. Aos poucos, a área delimitada pelo câmbio vai se tornando cada vez mais cilíndrica.
O câmbio vascular o caule, como o da raiz, também é um meristema de origem mista, primária e secundária. Isso porque tem origem tanto do cambio fascicular, um meristema primário, quanto do câmbio interfascicular, um meristema que surgiu da desdiferenciação de células parenquimáticas.
Câmbio suberógeno ou felogênio
A atividade do câmbio vascular faz com que o caule vá progressivamente aumentando de espessura. Para acompanhar esse crescimento em diâmetro, célula do parênquima cortical sofrem desdiferenciação e originam um cilindro de meristema secundário, o felogênio. Como vimos, a atividade do felogênio produz feloderme para o interior e súber para o exterior, formando a periderme, que passa a revestir o caule.
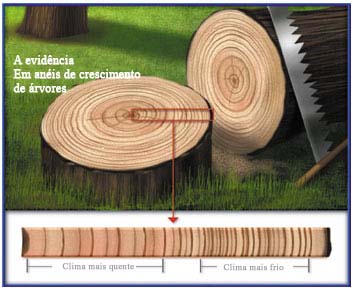
Anéis anuais
Nas regiões de clima temperado, a atividade do câmbio varia no decorrer do ano. A atividade cambial é muito intensa durante a primavera e o verão, diminuindo progressivamente no outono até cessar por completo no inverno. No fim do verão, quando está encerrando mais um ciclo de atividade, o câmbio produz vasos lenhosos de paredes grossas e lúmen estreito, que constituem o lenho estival. Na primavera, ao retomar o seu funcionamento depois do repouso invernal, o câmbio produz vasos lenhosos de paredes delgadas e lúmen grande, que constituem o lenho primaveril.
Troncos de árvores que vivem em regiões temperadas apresentam, portanto, anéis de lenho primaveril. Quando esses troncos são observados em corte transversal, esses anéis são facilmente identificáveis e o número de pares de anéis corresponde à idade da árvore.
Crescimento e desenvolvimento
O crescimento de uma planta começa a partir da germinação da semente. A hidratação da semente, por exemplo, ativa o embrião. As reservas contidas no endosperma ou nos cotilédones são hidrolisadas por ação enzimática. As células embrionárias recebem os nutrientes necessários, o metabolismo aumenta e são iniciadas as divisões celulares que conduzirão ao crescimento.
A radícula é a primeira estrutura a imergir; a seguir, exterioriza-se o caulículo e a plântula inicia um longo processo que culminará no vegetal adulto.
Qual a diferença entre crescimento e desenvolvimento?
Esses dois termos são frequentemente utilizados como sinônimos. No entanto, há uma diferença entre eles:
- O crescimento corresponde a um crescimento irreversível no tamanho de um vegetal, e se dá a partir do acréscimo de células resultantes das divisões mitóticas, além do tamanho individual de cada célula. De modo geral, o crescimento também envolve aumento do volume e da massa do vegetal. O crescimento envolve parâmetros quantitativos mensuráveis (tamanho, massa e volume).
- O desenvolvimento consiste no surgimento dos diferentes tipos celulares e dos diversos tecidos componentes dos órgão vegetais. É certamente um fenômeno relacionado ao processo de diferenciação celular. O desenvolvimento envolve aspectos quantitativos, relacionados ao aumento da complexidade do vegetal.
A ocorrência desses dois processos é simultânea. Um vegetal cresce e se desenvolve ao mesmo tempo
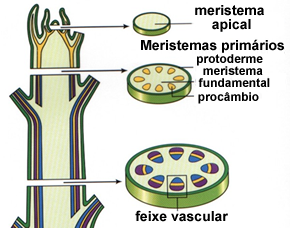
O meristema
Todos os tipos de células que compõe uma planta tiveram origem a partir de tecidos meristemáticos, formados por células que têm uma parede primária fina, pequenos vacúolos e grande capacidade de realiza mitose.
As células dos tecidos diferenciados, ainda que não tenham morrido durante a diferenciação (como o súber, o xilema, por exemplo), perdem a capacidade de se multiplicar por mitose. As células meristemáticas se multiplicam e se diferenciam, originando os diversos tecidos permanentes da planta, cujas células não mais se dividem.
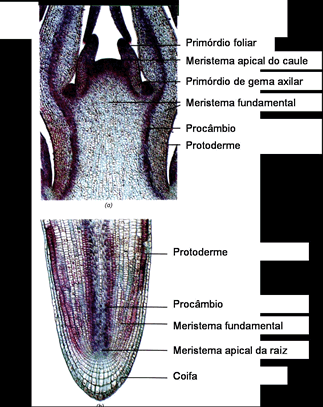
Meristema primários
Em certos locais das plantas, como nos ápices da raiz e do caule, existem tecidos meristemáticos que descendem diretamente das primeiras células embrionários, presentes na semente. Esses são os meristemas primários.
O caule cresce em comprimento graças à atividade de um meristema primário presente em seu ápice, o meristema apical caulinar. Já o meristema responsável pelo crescimento em comprimento da raiz não é terminal, mas está protegido sob um capuz celular chamado de coifa. Por isso é denominadomeristema subapical radicular.
| |  |
Meristemas secundários
Meristemas secundários são os que surgem a partir de células diferenciadas, geralmente parenquimáticas, que readquirem a capacidade mitótica, fenômeno que os botânicos denominam desdiferenciação. O felogênio que constitui a periderme, por exemplo, é um exemplo de meristema secundário, que surge pela desdiferenciação de células do parênquima localizadas sob a epiderme. A multiplicação das células do felogênio origina a feloderme e o súber que compõem a periderme.
Continue Lendo no blog:
http://www.sobiologia.com.br

Outro Link Para Estudo:
http://aulasdebotanica.blogspot.com.br/
Função
A função da flor é mediar a união dos esporos masculino (micrósporo) e feminino (megásporo) num processo denominado
polinização. Muitas flores dependem do
vento para transportar o pólen entre flores da mesma espécie. Outras dependem de
animais (especialmente insetos) para realizar este feito. O período de tempo deste processo (até que a flor esteja totalmente expandida e funcional) é chamado anthesis. A maior flor encontrada é a
Rafflesia arnoldii, espécie na qual alguns dos exemplares encontrados já chegaram a 1 metro de diâmetro e 11 kg.
[13]
Muitas das coisas na natureza desenvolveram-se para atrair animais polinizadores. Os movimentos do agente polinizador contribuem para a oportunidade de recombinação genética com uma
populaçãodispersa de plantas. Flores como essas são chamadas de entomófilas (literalmente: amantes de insetos). Flores normalmente têm nectários em várias partes para atrair esses animais.
Abelhas e
pássaros são polinizadores comuns: ambos têm
visão colorida, assim escolhendo flores de coloração atrativa. Algumas flores têm padrões, chamados guias de néctar, que são evidentes na
espectro ultravioleta, visível para abelhas, mas não para os humanos. Flores também atraem os polinizadores pelo aroma. A posição dos estames assegura que os grãos de pólen sejam transferidos para o corpo do polinizador. Ao coletar néctar de várias flores da mesma espécie, o polinizador transfere o pólen entre as mesmas.
O aroma das flores nem sempre é agradável ao nosso
olfato pode ser veneno fatal para as pessoas. Algumas plantas como a Rafflesia, e a PawPaw Norte-Americana (Asimina triloba) são polinizadas por
moscas, e produzem um cheiro de carne apodrecida para atrair esses ajudantes.
Outras flores são polinizadas pelo vento (as
gramíneas por exemplo) e não precisam atrair agentes polinizadores, tendendo assim a possuir aromas discretos. Flores polinizadas pelo vento são chamadas de anemófilas. Sendo assim o pólen de flores entomófilas costuma ser grudento e de uma granulatura maior, contendo ainda uma porção significante de
proteína (outra recompensa para os polinizadores). Flores anemófilas são normalmente de granulatura menor, muito leves e de pequeno valor nutricional para os insetos.
Existe muita contradição sobre a responsabilidade das flores nas
alergias. Por exemplo, o entomófilo Goldenrod(Solidago) é frequentemente culpado por alergias respiratórias, o que não é verdade, pois seu pólen não é carregado pelo ar. Por outro lado, a alergia é normalmente causada pelo pólen da anemófila Ragweed(Ambrosia), que pode vagar com o vento por vários quilômetros.
[editar]
Hermafroditismo
Ao contrário do que normalmente é lido, nenhuma flor pode ser considerada hermafrodita. Como hermafrodita, considera-se o organismo capaz de produzir
gametas masculinos e femininos. No entanto, a flor, por ser uma estrutura do
esporófito, é estritamente assexuada; não produz gametas e sim
esporos. Os esporos são responsáveis pela reprodução assexuada do vegetal. Dessa forma, a flor fica impedida de ser designada hermafrodita. A confusão deve-se à prática botânica que convencionou chamar o
megásporo de "esporo feminino" e o micrósporo de "esporo masculino", devido à diferença de tamanho entre eles - o mesmo parâmetro usado para diferenciar os gametas feminino (maior) e masculino (menor).

A flor é o aparelho reprodutor dos vegetais mais evoluídos, que são as Gimnospermas e as Angiospermas.
Nas Gimnospermas, que surgiram primeiro, as flores são incompletas, pois além de não possuirem sépalas e pétalas, que formam o revestimento externo, não produzem os óvulos dentro de um ovário, não formando, portanto, frutos.
As Angiospermas, por outro lado, surgiram mais tarde e suas flores são completas, nelas os óvulos, que irão produzir as sementes, se formam dentro de uma estrutura fechada, o ovário, que irá se transformar no fruto.
Origem das flores
As flores se originam de gemas ou botões florais localizados em posições diversas, geralmente nas extremidades dos ramos.
Constituição de uma flor completa
Pedúnculo - é ramo de caule em cuja extremidade a flor se forma
Receptáculo - exctremidade do pedúnculo onde as peças da flor se fixam
Sépalas - folhas modificadas, geralmente verdes, cuja função é de proteção. As sépalas fecham botão floral antes que este se abra. Seu conjunto denomina-se cálice
Pétalas - folhas modificadas, geralmente coloridas, cuja função é proteger os órgãos reprodutores e atrair pássaros ou insetos, que irão transportar os grãos de pólem de uma flor a outra. Seu conjunto forma a corola.
Estames - folhas modificadas que são os órgãos reprodutores masculinos do vegetal e cuja função é produzir os grãos de pólen. O conjunto dos estames é chamado de androceu
Carpelos - órgãos reprodutores femininos que formam um ovário onde serão produzidos os óvulos.O conjunto dos carpelos forma o gineceu ou pistilo.
Obs: o cálice, a corola, o androceu e o gineceu, que são os conjuntos formados por peças iguais, são denominados de verticilos florais.
Simetria Floral
Conforme a sua simetria as flores podem ser classificadas em:
Zigomorfas - com simetria bilateral
Actinomorfas - com simetria radial
Assimétricas - sem qualquer simetria
Verticilos protetores
O cálice e a corola são os verticilos protetores das flores e o seu conjunto pode ser chamdo de perianto (quando as sépalas e as pétalas são diferentes) ou perigônio (quando as sépalas e as pétalas são iguais em cor e tamanho). Neste último caso as sépalas e as pétalas são chamadas de tépalas.
| Aclamídea | sem cálice nem corola |
| Monoclamídea | apenas um dos dois, ou cálice ou corola |
| Diclamídea | Homeoclamídea | com cálice e corola iguais ou perigônio |
| Heteroclamídea | com cálice e corola diferentes ou perigônio |
Número de peças por verticilo
O número de peças por verticilo floral é uma das principais caraacterísticas usadas na classificação das Angiospermas.Assim, poderemos ter:
Flores dímeras - aquelas que têm duas peças por verticilo.

Flores trímeras - diz-se da flor organizada com base no número três, ou seis.

Flores tetrâmeras - diz-e da flor cujos verticilos se compõem de quatro elementos.

Flores pentâmeras - diz-se da flor organizada com base no número cinco ou múltiplo.

Verticilos reprodutores
Os verticilos reprodutores são o Androceu e o Gineceu, formados respectivamente por estames e carpelos.
Conforme possuam um ou os dois sexos, as flores podem ser: Monóclinas (hermafroditas) quando possuem os dois sexos, ou Díclinas quando os sexos são encontrados em flores diferentes.

Vegetais Monóicos e Dióicos
Os vegetais são monóicos ou hermafroditas quando os dois sexos são encontrados num mesmo indivíduo, numa mesma fllor (monóclina) ou em flores diferentes (díclinas).
Os vegetais dióicos ou unissexuados, por outro lado, possuem apenas um sexo em cada pé e suas flores, portanto, só podem ser díclinas.

Verticilos reprodutores
Androceu
É o verticilo reprodutor masculino, formado por folhas modificadas denominadas estames, cuja função é a produção dos grãos de pólen e que são constituidos por três partes:
| Antera - parte superior do estame com forma globulosa e cuja função é de produzir os grãos de pólen. |  |
| Conetivo - local por onde o filete se fixa na parte superior do estame que é a antera. |
| Filete- é a haste que liga o estame ao receptáculo da flor. |
Conforme o número de estames o androceu pode ser:
Isostêmone - quando o número de estames é igula ao número de pétalas
Anisosêmone - quando o nmero de estames é diferente do número de pétalas
Gamostêmone - quando os estâmes estão fundidos uns com os outros
Dialistêmone - com os estames estão livres, não fundidos
Antera
A antera é parte fertil do estâme onde serão produzidos os grãos de pólen.
Vamos analizá-la ainda jóven, antes de produzirem grãos de pólen e depois de madura, quando os grãos de pólen já estão formados.
Antera jovem - logo que a flor se abre a antera ainda não está madura e ainda não produziu grãos de pólen.
Fazendo-se um corte transversal, verifica-se que é dividida simetricamente em duas metades (tecas), existindo em cada uma delas duas cavidades denominadas sacos polínicos, revestidas por uma camada nutritiva de células denominada "tapetum". Verifica-se, também que, abaixo da epiderme, existe uma camada de celulas com paredes reforçadas, denominada endotécio ou camada mecância. Nesta fase, dentro de cada saco polínico são encontradas células que, como todas as demais, são diplóides, chamadas de "células mãe de grão de pólen".

Antera adulta - nesta fase as células-mãe, que eram diplóides, sofreram meiose e deram origem a grãos de pólen, haplóides e binucleados. Verifica-se que a camada nutritiva que rodeava os sacos polínicos desapareceu, bem como a camada intermediária (entre os sacos polínicos), ficando em casa teca uma grande cavidade, denominada câmara polínica, cheia de grãos de pólen. A partir deste momento a antera irá se abrir e o pólen será transportado até os órgãos femininos desta mesma flor ou de outra.
O grão de pólen
O grão de pólen é uma célula binucleada (núcleo vegetativo e núcleo reprodutivo) cercada por duas membranas a exina (mais externa) e a intina ( mais interna). A exina é cheia de rugas que permitirão a fixação na parte superior do carpelo, para que depois possa haver a fecundação.
O grão de pólen é um esporo (micrósporo) e ainda não é o gameta masculino do vegetal, este será formado mais tarde quando, dentro do tubo polínico o núcleo reprodutivo se dividir formando dois anterozóides.
Gineceu ou pistilo
É o verticilo reprodutor feminino das angiospermas, formado por folhs modificadsa, denominadas carpelos.
Os carpelos, que são folhas modificadas com os bordos fundidos, são constituidos de três partes:
| Estígma - parte suprior do carpelo, rugosa, onde deve cair o grão de pólen |  |
| Estilete - tubo que liga o estilete ao ovário, por onde deve crescer o tubo polínico |
| Ovário - parte basal, dilatada, onde são produzidos os óvulos, que formarão sementes |
Tipos de gineceu
Simples - formado por um só carpelo
Apocárpico - formado por diversos carpelos não fundidos. Dá origem a diversos frutos em uma mesma flor
Sincárpico - formado por diversos carpelos fundidos que vão formar um único fruto.
Tipos de ovário
Unicarpelar, unilocular - formado por um só carpelo que forma uma só cavidade (lóculo). É encontrado nos gineceus simples e nos apocárpicos.
Pluricarpelar unilocular - formados por diversos carpelos que formam uma única cavidade. É encontrado nos gineceus sincárpicos e forma frutos com uma única cavidade interna.
Pluricarpelar plurilocular - formado por diversos carpelos que formam diversas cavidades. É encontrado nos gineceus sincárpicos e forma frutos com o interior dividido em gomos.
Placentação
É o estudo da posição em que os óvulos se fixam na parede interna do ovário e, quando este se transformar no fruto, será a posição em que ficarão as sementes.
Tipos de placentação
Central - quando o óvulo se fixa no centro e no fundo do ovário. Ex: pêssego, manga, côco
Parietal - quando o óvulo se fixa nas paredes laterais do ovário. Ex: vagem, amendoim
Axial - nos ovários pluricarpelares pluriloculares, quando os óvulos se fixam no eixo central formado pelo ponto de encontro dos carpelos. Ex: laranja
Difusa - espalhada por toda a parede do ovário. Ex: mamão
O Óvulo
O óvulo, que é uma das principais conquistas evolutivas do Reino Vegetal , surgiu com o aparecimento da flor.
Além de produzir um gameta feminino dentro de uma estrutura protetora, ao se transformar na semente, cria um ambiente protegido onde o embrião pode sobreviver durante longos períodos esperando condições favoráveis para a germinação. E, quando esta acontece, proporciona as reservas nutritivas necessárias para que a plântula possa iniciar o seu crescimento.
Formação do óvulo
Nas Angiospermas, o óvulo ou os óvulos, começam a se formar no interior do ovário, a partir da placenta.
1 - No início é um pequeno calombo dentro do qual há uma célula grande, diplóide, denominada "célula mãe de megasporo"
2 - A célula mãe sofre meiose e forma quatro megasporos haplóides
3 - Dos quatro megasporos três degeneram e sobra apenas um.
4 - O núcleo do megasporo se divide por mitose formando oito núcleos haplóides
5 - Os oito núcleos originam as sete células que formam o saco embrionário: a oosfera, as sinérgides, as antipodas e uma grande célula central - o mesocisto - com dois núcleos (núcleos polares).

Veja a descrição do óvulo abaixo:
| Tegumento - ou casca, formado por duas camadas de células: a primina(mais externa) e a secundina (interna). |  |
| Nucela - tecido nutrivo do óvulo das Angiospermas que envolve o saco embrionário. |
| Saco embrionário - formado pela meiose de uma célula mãe inicial e composto, em muitos casos, por sete células haplóides: a oosfera, duas sinérgides, três antipodas e uma grande célula central, o mesocisto, com dois núcleos (núcleos polares). É a parte fértil do óvulo. |
| Funículo - é o pedúnculo que liga o óvulo (e mais tarde a semente) à placenta. |
| Hilo - porção terminal do funículo. É o lugar por onde, mais tarde, a semente se destacará do fruto. |
| Chalaza - parte basal da nucela que se liga ao funículo através do hilo. |
| Micrópila - pequena abertura na extremidadade superior do óvulo. Geralmente é por ela que penetra o tubo polínico. |
| Placenta - local na parede do ovário onde o óvulo se fixa e por onde, mais tarde, a semente se liga ao fruto.. |
Tipos de óvulo
Ortótropo - é o óvulo em que o hilo a chalaza e a micrópila estão em uma mesma lina reta.
Anátropo - é o óvulo cujo eixo principal é curvo e a micrópila fica virada para a placenta.
Campilótropo - é o óvulo recurvado em que a micrópila e a chalaza se aproximam em um mesmo plano horizontal.

Tipos de flores em relação à posição do ovário
Hipógina - é a flor em que o ovário se coloca no receptáculo em um ponto que fica acima do plano em que as outras peças se fixam.
Perígina - é a flor em que as demais peças se fixam em um plano que corta o ovário.
Epígina - é a flor em que o ovário é envolvido pelo receptáculo ficando todo ele abaixo do plano em que as outras peças se fixam.
fonte :http://www.universitario.com.br/celo/topicos/subtopicos/botanica/anatomia_vegetal/flor/flor.html